 Шамамен 318 миллион жыл бұрын, Пермь кезеңінде біздің планетамызда биологиялық синапсидтер мекендеген. Бұл ежелгі тіршілік иелері қазіргі бауырымен жорғалаушылардан тек мөлшерімен ғана емес, сонымен қатар стоматологиялық құрылымымен де ерекшеленді. Өткізгіштерден басқа, олардың жыртқыштық сипатын көрсететін шоқтары да болды. Пермь шөгінділерінде жақсы сақталған көптеген қазба қалдықтарын зерттеу негізінде ғалымдар осындай батыл тұжырымдар жасай алды. Кейіннен, бұл тарихқа дейінгі тіршілік иелері диметродонттар деп аталды.
Шамамен 318 миллион жыл бұрын, Пермь кезеңінде біздің планетамызда биологиялық синапсидтер мекендеген. Бұл ежелгі тіршілік иелері қазіргі бауырымен жорғалаушылардан тек мөлшерімен ғана емес, сонымен қатар стоматологиялық құрылымымен де ерекшеленді. Өткізгіштерден басқа, олардың жыртқыштық сипатын көрсететін шоқтары да болды. Пермь шөгінділерінде жақсы сақталған көптеген қазба қалдықтарын зерттеу негізінде ғалымдар осындай батыл тұжырымдар жасай алды. Кейіннен, бұл тарихқа дейінгі тіршілік иелері диметродонттар деп аталды.
Олар ұзындығы 3,5 метрден асатын қатты денелі аңдар еді. Олардың айрықша ерекшелігі - дорсалды парус деп аталады. Бұл бүкіл омыртқа бойымен жүретін жоғары тері қатпарлары еді. Ұқсас түрдегі өсінділер ежелгі амфибиялар мен жануарлардың кейбір түрлерінде байқалды, олардың қатарына динозаврлар мен пеликозаврлар да кіреді. Мүмкін, парус осы жануарлардың денесін терморегуляция процесіне белсенді қатысты. Егер сол күндері қоршаған ортаның температурасы өте жоғары болғанын ескерсек, онда суық қанды диметродон, егер парус болмаса, қызып кетуден оңай өліп кетуі мүмкін. Сонымен қатар, мұндай түпнұсқалық терінің пайда болуы жыныстық кезеңнің барысында диметродон қолданатын қайталама жыныстық атрибут рөлін атқара алады, сонымен қатар ол тығыз тропикалық таулы жерлерде болған кезде оны жасыру үшін қолдана алады. Басқа болжам бойынша, кейіпкеріміз терінің қатпарларын жүзу кезінде парус ретінде қолдана алады.
Өмір салтына келетін болсақ, диметродондар шағын топтарда өмір сүрді. Ересектер саванналарды, ал жас адамдар жаңбыр ормандары көп болған жерлерді көрді. Бірақ шөлде диметродон өмір сүре алмады. Мұндай ыстық ауа-райы оған ұнамады.
Бұл жануарлар әлемінің барлық өкілдеріне шабуыл жасаған қатыгез және қатал жыртқыш еді. Ұстара өткір тістерімен және күшті жақтарымен ол жәбірленушінің денесін оңай бөліп тастады.
Құрылымы мен өмір салтында рептилияларға қарағанда сүтқоректілерге көбірек ұқсайтын. Ғалымдар оны ірі өкілі болған пеликозаврлардың отрядыға апарды.
Диметродондардың қандай себеппен жойылып кеткендігін ғалымдар нақты білмейді. Мүмкін бұл күрт климаттық өзгерістерге байланысты болса керек, суық қанды жануарлар өмір сүре алмады. Тағы бір болжам бойынша, дамыған тіршілік иелері оларды еліктірді.
Ерте оқыту
Диметродонның қалдықтары алғашында 1870 жылдары Эдвард Сусыншы Коуппен суреттелген. Ол оларды Техастағы Қызыл Градтардың пайда болуынан алынған басқа Пермь тетраподтарының жиынтығымен бірге қабылдады. Коллектор Джейкобс Бол, геолог У.Ф.Кумминс және палеонтолог Чарльз Стернберг оларды еңсеруге тапсырды.Осы үлгілердің көпшілігі қазір Американың табиғи тарих мұражайында немесе Чикаго университетінің Уокер музейінде.
Штернберг өзінің бірнеше үлгілерін Мюнхен университетінің неміс палеонтологы Фердинанд Бройлиге жіберді, бірақ ол оларды да, Копты да зерттемеді. Эдвардтың қарсыласы Чарльз Марш сонымен бірге диметродонның бірнеше сүйегін жинады, бірақ ол оларды Уокер мұражайына берді.
Аты Диметродон 1878 жылы қолданылған күрес, үш түрді бөліп көрсетті - D. incisivus, D. тік ішек және D. гигастар.
Диметродонмен әйгілі суреттердің бірі
Алайда, диметродон қалдықтарының алғашқы сипаттамасы 1875 жылы, Коп сырғанаптарды сипаттаған кезде пайда болды C. limbatus. Бұл жануар диметродонмен бірдей аймақта табылды, ал 19-шы ғасырдың аяғы мен 20-ғасырдың басында пеликозаврлардың көп бөлігі диметродонға немесе клепсидоптарға жатады. 1940 жылы бұл туралы жарияланым жарияланды C. limbatus бұл диметродонның бір түрі.
Желкенмен сипатталатын алғашқы синапс - бұл ақылды адамдар. C. natalis, сонымен қатар Cope атап көрсетті. Ол желкенді фин деп санады және оны базиск кескінінің шоқымен салыстырды. Желкен D. incisivus және D. гигастар консервіленген жоқ, алайда, үлгі D. тік ішек ұзартылған жүйке омыртқалары. Алайда, 1886 жылы Коп парустың мақсатын елестету қиын деп түсіндірді. Оның айтуынша, егер жануар су өмір салтын жүргізбесе, онда парус немесе аққулар қозғалысқа кедергі келтіреді, ал аяқтар мен тырнақтар басжилиск сияқты арборальды өмір салтын жүргізуге жеткілікті үлкен болмады.
ХХ ғасыр
ХХ ғасырдың басында Э. Кейс бірнеше жаңа түрлерді бөліп, диметродон туралы байыпты зерттеу жүргізді. Оған Карнеги институты қаржылай көмек көрсетіп, Америкадағы түрлі мұражайлардың материалдарын ұсынды. Көптеген үлгілерді бүкіл ұрпақты тек фрагменттермен сипаттаумен әйгілі болған Коп сипаттаған, бірақ ол бұл қалдықтарға көп мән берген жоқ.
1920 жылдардың аяғында Альфред Ромер көптеген демитродон үлгілерін қайта қарап, тағы бірнеше түрді бөліп көрсетті. 1940 жылы Ромер мен Ллевеллин Прайс Cope сипаттаған синапсидтердің көпшілігін қарастырған Pelicosaurus шолуын өткізді. Осы зерттеу нәтижелерінің көпшілігі әлі күнге дейін өзекті.
Dimetrodon incisivus түрлерінің ескі қалпына келуі
Ромер мен Прайс жарияланғаннан кейін Оклахома мен Техастың сыртындағы диметродонның көптеген үлгілері табылды. Сонымен, 1966 жылы Юта штатында кішкене сынықтар табылды, ал 1969 жылы Аризонада қалдықтар табылды. 1975 жылы Олсон Огайо штатында диметродонның ашылғанын хабарлады. 1977 жылы Берман Нью-Мексикодағы материалдар негізінде түрді сипаттады. D. occidentalis («батыс»), оған Юта мен Аризонаның қалдықтары да енген.
Осы ашылуларға дейін Техас пен Оклахома қалған аумақтардан Орта-континентальды теңіз жолы арқылы бөлінді деп есептелген, сол себепті кіші сфенакодон Солтүстік Американың батысында мекендеген. Жаңа табылулар, олар ішкі теңіздің болуын жоққа шығармайды, бірақ оның шектеулі табиғатын және оның диметродонды қоныстандыруға кедергі болмағандығын көрсетеді.
Сипаттамасы
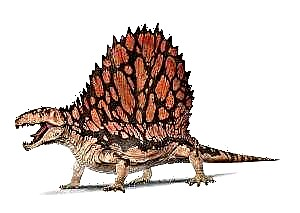
Диметродонның өте үлкен басы бар, оның иықтары өткір тістері бар. Ол белсенді аңшы болған шығар: ол қосмекенділерді, бауырымен жорғалаушылар мен балықтарды аулаған. Алдыңғы тістерімен диметродон құрбанды ұстап, оны жыртып тастады. Артқы тістері артқа бүгіліп, олардың көмегімен жануар ұсақ тіршілік иелеріне кесіліп, үлкен ет кесектерін шайнады. Диметродонның денесі бөшке тәрізді болды. Диметродонның ең керемет ерекшелігі - бұл тері астындағы парус омыртқалардың өсінділерінен асып түседі. Осыған ұқсас желкендер әртүрлі тарихқа дейінгі жануарларда дамыған (Platigistrix amphibians, Edaphosaurus and Secodontosaurus, Spinosaurus динозавр) және температура реттегіші ретінде қызмет етті. Басқа нұсқаларға сәйкес, желкенді отырыс ойындарында пайдаланған, өсімдіктердің тік сабақтарының арасында камуфляж ретінде қызмет еткен немесе жүзген кезде нағыз желкенді. «Парус» өмір бойы біртіндеп дамып келеді, төмен спинді процестермен дамитын диметродонның жас тұлғалары табылды. Жас адамдар, су қоймаларының жағасында, ересектер сияқты таулы жерлерде өмір сүрген сияқты. Әйтсе де, шөлге қарсы танымал кітаптардағы диметродонның бейнесі шындыққа жанаспайтын болып көрінеді - бауырымен жорғалаушы болмай, ол ыстық, құрғақ жерде өмір сүре алмады.
Бас сүйегі
Диметродонның бас сүйегі жоғары, кейіннен сығылған. Премаксила жақ сүйегінен терең диастемамен бөлінеді. Алдыңғы шеті қатты дөңес. Кішкентай танаулар мұрынның алдында орналасқан. Жыртылған сүйек мұрынға жетпейді. Көз розеткалары бас сүйегінің артқы жағында орналасқан. Премаксилада үш тіс бар, жоғарғы жақтың екінші тісі - канин тәрізді, үлкейген, иілген, жиектері қатайған. Тістері өте жұқа, альбертозавр тәрізді. Олардың негіздерінде кішкентай қуыстар болды, бұл тістерге жүктемені азайтады, бірақ оларды тістердің зақымдануынан сақтамайды. Ат D. тетонис ешқандай сыну болған жоқ, дегенмен жиектері өте өткір болды. Жамбас маңындағы екі жақтың тістері азаяды, олардың артында олар біртіндеп азаяды. Формасы бойынша диметродонның тістері мен оның туыстары тамшыларға ұқсайды, бұл сфенакотамусты басқа ерте синапсидтерден ажыратуға көмектеседі.
2014 жылғы зерттеу көрсеткендей, диметродондар қару жарысының өзіндік түрін өткізді. Шағын D. миллери олар кішкентай олжаны аулағандықтан, тістерін сындырмады. Түрлердің әртүрлілігі мен мөлшері ұлғайған сайын тістер пішінін өзгертті. Ат D. лимбатус тістердің секодонтозавр тәрізді кескіш пішіні болды. Үлкен көлемде D. әжесі тістер акулалар мен терроподтар тәрізді болды. Осылайша, диметродондар дамып келе жатқанда мөлшерін ұлғайтып қана қоймай, сонымен бірге аң аулаудың әдеттегі құрылғыларын да өзгертті.
Диметродон завропсидтен фенестраның қатысуымен ерекшеленеді. Рептиляторлардың екі фенестры бар немесе олар мүлде жоқ, ал синапсидтерде осындай тесік бар еді. Диметродонда ерте тетроподтардан сүтқоректілерге төменгі жақтың артқы жоталары мен мұрын қуысы тәрізді ауысулардың ерекше белгілері болған.
Мұрын қуысының ішкі жағында арнайы жоталар, турбиналар орналасқан. Олар иісті эпителийдің ауданын ұлғайтып, шеміршекке қолдау көрсете алар еді. Бұл жоталар сүтқоректілерге және одан кейінгі синапсидтерге қарағанда кішкентай, оларда назотурбиндер жылы қандылықтың белгісі болып табылады. Олар шырышты қабықты жылытып, кіретін ауаны ылғалдандыруы мүмкін. Осылайша, диметродон ішінара жылы қанды жануар болды.
Диметродонның тағы бір ерекшелігі - шағылысқан пластина деп аталатын жақ сүйегінің артындағы доғалдау. Ол төртбұрышты сүйекке жалғанған сүйек үстінде орналасқан, бірге жақ сүйегін біріктіреді. Кейінгі синапсидтерде ортаңғы құлақтың сүйектері - мальлюска түзетін жақ сүйектерінің буынынан ажыратылған артикулярлы және шаршы сүйектерінің процестері. шағылысқан пластина кейіннен қазіргі сүтқоректілердің құлақ сүйектерін қолдайтын тимпаникалық сақинаға айналды.
Құйрық
Ұзақ уақыт бойы диметродон қысқа құйрығы бар жануар ретінде ұсынылды, өйткені денеге жақын 11 құйрықты омыртқалар белгілі болды, олар жамбас сүйектерінен алыстаған кезде қатты тарылып, алғашқы сипатталған күйінде құйрық мүлде жоқ немесе жақсы күйде болмаған. Тек 1927 жылы 50 омыртқадан тұратын диметродонның толық құйрығы табылды. Ол дененің ұзындығының көп бөлігін есептеді және қозғалу кезінде тепе-теңдік қызметін атқарды.
Желкен
Қаңқа D. лумизи
Диметродонның ең жарқын және көрнекті белгілерінің бірі - бұл жұлын және мойын омыртқаларының жоғары спиналық процестері. Тұқым ашылған кезден бастап олар әртүрлі жолмен суреттелген: жай шыбықтарды, сондай-ақ былғарыдан, тіпті бүркіттен тұратын «парус» сияқты. Парус бір метр биіктікке жетті. Нейрондық омыртқалардың жоғарғы жағы мүйізді жамылғылармен қапталған. Әрбір нейрондық шыбықтың ерекше формасы бар, бұл дифференциация тіпті «диметродонтовая» деп аталады. Шыбықтар омыртқалы дененің жанында тікбұрышты пішінге ие, олар одан алыстаған сайын сегіз пішінді алады. Бұл форма процестерді күшейтіп, сынықтарға кедергі келтіреді деп саналады. Бір жеке тұлға белгілі D. giganhomogenes толығымен тікбұрышты жүйкелік омыртқалармен, алайда орталықтың жанында «сегіздіктің» іздері бар. Сірә, бұл өзгерістер адамның жасына байланысты. Әр шыбықтың микроанатомиясы бұлшықеттерге бекітілген жерді және парустың өту орнын көруге мүмкіндік береді. Шоқтың төменгі, проксимальды бөлігі тегіс беткейге ие. Мүмкін, оған эпаксиалды және гипаксиялық бұлшықеттер, сонымен қатар өткір талшықтар деп аталатын дәнекер тінінің желісі қосылған болуы мүмкін. Омыртқалардың дистальды бөлігі тегіс, бірақ периостумға көптеген ойықтар енеді, мүмкін олардың өмірінде қан тамырлары болған. Нейрондық омыртқалардың көлденең қимасының көп бөлігін құрайтын көп қабатты ламинатталған сүйекте көптеген өсу сызықтары бар, олардың көмегімен сіз өлген кезде әр адамның жасын анықтай аласыз.
Ерекше ойық барлық омыртқалардан өтеді. Бұрын бұл жерде қан тамырлары орналасқан деп есептелген, бірақ сүйектерде олардың іздері болмағандықтан, ойық басқа нәрсеге арналған болуы мүмкін, ал желкендегі кемелер саны ойлағандай аз болған.
Диметродон қаңқасын заманауи қайта құру. Жариялаған Скотт Хартман
Патологияны зерттеу кейбір нейрондық омыртқалардың сынғанын, содан кейін сауыққанын көрсетті. Бұл процестердің кем дегенде бір бөлігі параллельмен (немесе ұқсас тінмен) жабылған, бұл оларды зақымданғаннан кейін орнында ұстап, нәтижесінде оларды сауықтыруға болатындығын білдіреді. Бірақ олар сонымен бірге шыбықтардың шыңдары көбінесе иіліп, кейде өте күшті болатындығын байқады, бұл шыбықтардың шыңдары параққа кірмегенін білдіреді. Осы «паруспен шығу» конфигурациясының одан әрі расталуы процестердің беткі құрылымынан болады. Олар артқы бұлшықеттерге байланған жерде өрескел болды, содан кейін олар тегіс бола бастады, дәл сол жерде парламента шыбықтары пайда болды. Сынықтар орнында өсетін кортикальды сүйек өте тамырлы, бұл паруста жұмсақ тіндердің және қан тамырларының белгілі бір санының болуын көрсетеді.
Диметродон терісінің іздері белгісіз, сондықтан кез-келген нақты құрылым туралы айту қиын. Кейінірек синапсидтер, мысалы, эсемменоз, тегіс терісі бар, көптеген бездері бар. Алайда, Асцендонан сияқты неғұрлым қарапайым ерте варанопеидтерде таразының іздерін анықтауға болады. Диметродонның дененің төменгі жағында қабыршақты құрылымы болуы мүмкін, ал оның бүйірлері мен үстінде терісі терапевтік агент сияқты тегіс болған.
Гаит
Диметродон дәстүрлі түрде жерді сүйретіп жатқан ішекті «кескелдирікпен» бейнеленген, алайда жақында диметродонға немесе оның жанындағы синапсидке жататын жолдар табылған, олар түзетілген аяғымен қозғалатын, асқазан мен құйрығын жерден мүлдем бос ұстайтын жануарды көрсетеді.
Әрине, диметродон қалаған кезде көңілді болуы мүмкін. Алайда, жүру және жүгіру кезінде оның аяқ-қолдары әлі де түзетілген күйге ие болды, осыған байланысты диметродон оның құрбандарына қарағанда тезірек болуы мүмкін (қосмекенділер мен кіші синапсидтер).
Белгілі түрлер
- D. тетонис Рейз және Берман, 2001. Жоғарғы қызыл төсектің төменгі қабаттары (вольфкамп), Германия, Бромакер және Ресей. Ең кішкентай диметрод, салмағы шамамен 24 кг. Солтүстік Америкадан тысқары белгілі белгілі түрлер. Биік парусқа ие болды. Ол жер биотасына бекітілді.
Диметродонның қаңқасы
- D. миллери Ромер 1937. Сакмара кезеңі, ұзындығы 174 см, Путнам формациясы, Техас. Екі қаңқамен белгілі: толық дерлік MCZ 1365 және үлкенірек, бірақ соншалықты жақсы сақталмаған MCZ 1367. Техас диметродонының ең алғашқы түрі. Нейрондық омыртқа құрылымында ол басқа түрлерден ерекшеленеді: D. миллери олар дөңгелек пішінді, ал басқа түрлерде олар сегізге ұқсайды. Бұл омыртқалар да басқаларға қарағанда қысқа. Бас сүйегі жоғары, мұрты қысқа. Осындай құрылым да бар D. booneorum, D. лимбатус және D. әжесімүмкін солай D. миллери олардың атасы болған. Жақын D. occidentalis. Синх.Clepsydrops natalis Күрес, 1887 жыл.
- D. natalis 1877 ж. Жеңілдігі, американдықтардың ең кіші түрі. Төмен трапециялы паруспен суреттелген, алайда, бұл формасы белгісіз. Бас сүйегінің ұзындығы шамамен 14 см, салмағы - 37 кг. Техас. Бас сүйегі төмен, үстіңгі жақта жалаңаш тістер бар. Ірі дененің жанында табылған D. лимбатус.
Қаңқа D. incisivus
- D. лимбатус Сакмара және Артинский деңгейлері - бас сүйегінің ұзындығы 40 см-ге дейін, жалпы ұзындығы 2,6 м-ге дейін, Техастағы Адмирал және Белл жазықтары. Паруспен танымал ең алғашқы синапсид. Әдебиетте жиі бейнеленген. Ол бастапқыда сипатталған Clepsydrops limbatus, оны 1940 жылы Ромер мен Прайс диметродонға жеткізді. Синх.Clepsydrops limbatus Күрес, 1877,? Dimetrodon incisivus Күрес, 1878 ж. Dimetrodon rectiformis Күрес, 1878 ж. Dimetrodon semiradicatus Cope, 1881.
- D. incisivus 1878 ж. Жеңілдігі - кейде типтік деп саналатын алғашқы түрлердің бірі. Мүмкін синоним D. лимбатус.
- D. booneorum Ромер 1937 - Артинский кезеңі - ұзындығы 2,2 метрге дейін, Техас. Ромер 1937 жылы сипаттаған.
- D. гигашомогендер 1907 іс. Кунгурск қабаты. Биіктігі 3,3 метрге жетті. Бас сүйегі қысқа және салыстырмалы түрде ұзын. Бабалардың бірі D. Ангеленсис. Arroyo Formation табылған.Case 1907 жылы қайта шақырылған, ол әлі де жарамды.
Қаңқасы D. grandis Case, 1907
- D. әжесі Іс, 1907. Ерте Кунгурский кезеңі. Оның ұзындығы 3,2 м-ге жетті, бас сүйегі төмен, ұзындығы 50 см.Оның тек төрт ісік алды тістері болды. Арройо формациясының қалыптасуында табылған, Техас. Синх. Theropleura grandis Іс, 1907 ж. Батиглиптус теодори Іс, 1911,? Диметродон гига Күрес, 1878 ж. Dimetrodon maximus Ромер, 1936,? Диметродон б.з.д. гига Grandis Штернберг, 1942.
- D. лумизи Ромер 1937. Кунгур деңгейі. 2,5 метрге дейін өсті. Arroyo Formation-де табылды, Техас. Онда бас сүйегінің төмен және қанатты парус пішіні бар.
- D. Ангеленсис 1962 ж. Ескі Уфа дәуірі (кеш Кунгур). Соңғы және ең танымал түрлер. Тіршілік кезінде ол 4-4,5 метрге дейін өсті. Техастағы Сан-Анджелода пайда болған. Бас сүйегі ұзын, 50 см-ге дейін, ал төмен, жоғарғы жақтары ұзын жіңішке. Барлық үлгілер нашар сақталған. Күнә :? Эосёдон худсоны Олсон, 1962 (номен дубий) ,? Дала гүлі Олсон және Бирбауэр, 1953.
- D. бореалис Лейди, 1854.270 миллион жыл бұрын, ханзада Эдвард аралы. Мүмкін болатын түрі, батигнат ретінде де белгілі. Осы аймақтағы қалдықтардың жасы өсімдік қоқысын зерттеуден кейін расталды. Кейінірек бигаттың бүкіл басы табылды. Бас сүйегінің ұзындығы небары 40-45 см.
Диметродонның ізі
- D. occidentalis Берман 1977 - Нью-Мексикодағы Abo / Cutler Formation-тен шыққан жалғыз диметродон. Оның ұзындығы 1,5 метрге жетуі керек. Бұл атау «Батыс Диметрод» дегенді білдіреді. Бір кішкентай қаңқамен белгілі. Шамамен байланысты D. миллери.
- D. гигастар Пермьдегі Артинский және Кунгурский деңгейлері. Ол бастапқыда сипатталған Гигаларды тазартатын клетчаткалардегенмен, кейінірек ол диметродон ретінде жіктелді. Жақсы сақталған бірнеше үлгілер түрге жатады. Сондай-ақ синоним деп саналады D. әжесі.
- D. макроспондилус Іс, 1907 - Коп былай деп сипаттады Macrospondylus клеткалары, диметродон үшін Case анықталады.
Не тамақтандырды
Диметродонның бас сүйегінің сүйектері өте жұқа болғанына қарамастан, оның күшті жақтары өткір тістермен кесілген, ол жәбірленушіге мықтап шағып алған. Тістері әртүрлі мөлшерде болды, молярлар артқа бүгілді. Алдыңғы ұзын тістерімен, қазіргі арыстандар сияқты, ол құрбандықты тістейді. Диметродон олжасын ұстап алды. Ғалымдар бұл жыртқыш қосмекенділерді, бауырымен жорғалаушылар мен балықтарды аулаған деп санайды. Алдыңғы тістерімен диметродон құрбанды ұстап, оны жыртты. Артқы тістері артқа қарай иіліп, олардың көмегімен жануар кішкене қылшықтарды жұлып алып, үлкен ет кесектерін шайнады.
ӨМІР САЛТЫ
Диметродон ерте Перм кезеңіндегі ең ірі және ең жыртқыштардың бірі болды. Бұл жануарлар жер бетінде алғашқы динозаврлар пайда болғанға дейін жоғалып кетті.
Диметродондардың қалдықтарын зерттеген палеонтологтар жақсы қаруланған және қатал жыртқыштар деген қорытындыға келді. Диметродон заманауи көліктің өлшемі болды. Оның қысқа күшті аяқ-қолдары болған, сондықтан ғалымдар диметродон заманауи кесірткелер сияқты қозғалады деп сенеді.
Ол өте баяу жануар болған шығар. Диметродонның қанша салмақ түсіргені туралы нақты мәліметтер жоқ, бірақ оның массасы айтарлықтай болған деп есептеледі. Артқы жағындағы үлкен, парус тәрізді өсім оған қорқынышты көрініс берді. Осы «парустың» көмегімен жануар денесінің температурасын реттей алады деп сенеді.
Таңертең күн суыған диметродон жылу парус арқылы осы динозаврдың денесінің басқа бөліктеріне жіберілді, сондықтан оны ықтимал құрбан болғандарды қыздырды. Салқындау үшін, диметродон парусын суға батыруға жеткілікті болды. Ғалымдар диметродондардың неліктен жоғалып кеткенін білмейді.
ЖАЛПЫ ЕРЕЖЕЛЕР. СИПАТТАМА
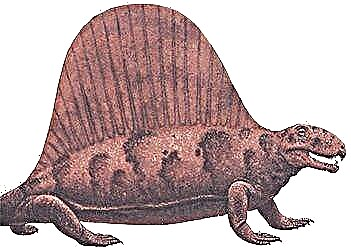
Пермь, 280 миллион жыл бұрын
Солтүстік Америка
Ұзындығы 3,5 м
Бұл пеликозаврлардың ішіндегі ең әйгілісі, ең көне жануарлар-кесірткелер. Денесі мықты, аяқтары қысқа, жақтары мықты, өткір тістері бар. Артқы жағында жоғары былғары парус орналасқан, оны омыртқалардың спиналық процестері қолдады. Оның функциялары нақты белгілі емес. Көптеген ғалымдардың пікірінше, «парус» дене температурасын оңтайлы ұстауға көмектесті: күнде желкеннің қан тамырларындағы қан қызып, көлеңкеде салқындатылды. Бұл сексуалдық серіктестерді тарту үшін ашық түсті тарақ болған шығар.
ҚЫЗЫҚТЫ АҚПАРАТ. ОСЫ БІЛСІҢІЗ.
- Диметродон атауы грек тілінен шыққан. Ол «димитро» екі сөзден тұрады - «екі өлшемде бар нәрсе» және «дон», яғни «тіс».
- Палеонтологтар диметродонның «парусының» қалдықтарын ұзақ зерттеді, соның негізінде парус осы қазбалы жануардың дене температурасын реттеу үшін пайдаланылды деген тұжырым жасалды.
- Диметродонның артқы жағындағы «парус» терісі қазіргі рептилиялар сияқты су өткізбейтін шығар.
Диметродонның сипаттамалық сипаттамалары
Парус: парус тәрізді өсу диметродонның артқы жағында мойыннан жамбасқа дейін болды Ғалымдардың пікірінше, желкен температураны бақылау жүйесінің өзіндік бір түрі болған. Таңертең диметродон күнді басады, күн сәулесі желкенді жылытады, сол арқылы жылу жануардың басқа мүшелеріне жіберілді. Мүмкін, қызып кетпес үшін, диметродон парусты суға батырған. Басқа нұсқа бойынша, парустың басқа функциялары болуы мүмкін, мысалы, жыныстық атрибут ретінде қызмет еткен - еркектер әйелдерге қарағанда үлкенірек және ашық парусқа ие бола алады.
Тістер: Ұзын, күшті көз тістері олжаны ұстап, оны бөліп тастау үшін қолданылған. Қысқа молярлар артқа бүгілді, олардың көмегімен диметроддон олжаны ұстап, етті жеп қойды.
Бас сүйегі: басы үлкен болды. Орбиталардың артында орналасқан тесік бас сүйегінің массасын азайтты. Бас сүйегінің артқы жағына күшті бұлшықеттер бекітілген.
Аяқтар: бұл жануардың артқы және алдыңғы жақтары қысқа және жаппай болды. Олар осы алып панголиннің дене салмағын қолдауға мәжбүр болды. Сонымен қатар, артқы аяқтардың күшті бұлшықеттері ұзын құйрықты ұстады.

- Археоптерикс қазбалары туралы мәліметтер
Қайда және қашан өмір сүрген
Қазіргі уақытта 6 археоптерикс қазбалары табылған. Бәрі Баварияда. Археоптерикс өмір сүрген күндерде Германия аумағы мүлдем басқа көрінетін және тропикте орналасқан континенттің бөлігі болды. Шөгінділер табылған тақталардың геологиялық жасын анықтауға сүйене отырып, Археоптерикстің жоғарғы юра кезеңінде, яғни шамамен 150 миллион жыл бұрын өмір сүргені белгілі болды.
Өмір сүру орны
Қазба қалдықтарының көп бөлігі Америка Құрама Штаттарында табылды, бірақ оларды Германияда да кездестіруге болады (Пермь кезеңінің басында бұл аумақтар құрлыққа біріктірілді). Диметродондар су объектілерінің қасында орналасқанымен, олардың тіршілік ету ортасы жеке адамдар жетілуімен өзгерді: жас жануарлар тығыз өсімдіктері бар батпақты жерлерді, жас ұрпақ көлдердің жағасын, ал тәжірибелі адамдар кең өзен аңғарларын таңдады. Мүмкін, диметродондар жартылай сулы өмір салтын ұстанған және жақсы жүзген шығар.
Көрініс ерекшеліктері
«Диметродон» атауы «екі тістің түрін» білдіреді. Кішкентай тістерден басқа, қабыршақтар мен қиғаштар жануарлардың жақтарында орналасқан (олардың саралануы сүтқоректілерге тән). Бауырымен жорғалаушылардан диметродон аяқ-қолдардың құрылымдық ерекшеліктерін иемденді, олар бүйірлерінде кең орналасып, денесінің астына тігінен емес, сонымен қатар суық қандылыққа ие болды. Оның дене температурасы қоршаған ортаға байланысты болды. Желкеннен басқа, жануарға тән қасиет, кем дегенде 50 омыртқадан тұратын өте ұзын құйрық болды. Диметродондардың мөлшері олардың түріне байланысты әртүрлі болуы мүмкін - дененің ұзындығы бойымен шашырау 0,6 ден 4,6 м дейін өзгерді.

Синапсидті бас сүйек уақытша қуыстарға белгілі. Олар екі жағынан бір-біріне, артында және орбиталардың астында орналасқан. Депрессиялар жақ бұлшықеттерін нығайтуға қызмет етті. Олардың болуы синапсидтердің тістенуін қосмекенділердің мүмкіндіктерімен салыстырғанда анағұрлым тиімді етті, бұл кезде бас сүйек құрылымының мұндай ерекшелігі болмаған.
Құрылымдық ерекшеліктері
Диметродонның омыртқалардың ұзын сүйек процестерінен тұратын дорсалды парусы болған, терімен қапталған. Ол терморегулятор қызметін атқара алады, күн тез қызады. Ғалымдар паруссыз ересектерге арналған диметродонның дене температурасы 3 сағат 40 минутта 6 ° -ке, ал онымен бірге 1 сағат 20 минутқа көтеріледі деп есептейді. Көлеңкеде былғары әшекейлер тез жылуды жіберіп, малды қызып кетуден сақтады. Сонымен қатар, желкенді жұптасу ойындары кезінде әйелдерді қызықтыру үшін пайдалануға болады (еркектерде мұндай доральды герб дамыған деп болжанады). Ол біртіндеп, диметродон жетіле бастаған кезде пайда болды.

Доминантты жыртқыш
Диметродон өз кезеңінің ең ірі жыртқышы болып саналады. Көрші жерде онымен бірге тұрған кез-келген жануарларды аулай алады. Диметродон иіс сезімі дамыған деп саналады. Бұл жануарларда жыныстық диморфизм көрінді, яғни аналық және еркектерде бастапқы жыныстық сипаттамаларға байланысты емес айырмашылықтар болды (мысалы, аналықтары аз болуы мүмкін). Диметродондардың нақты қалай өмір сүргені белгісіз: топтарда немесе жеке-жеке. Ер адамдар бір-біріне деген агрессияны көрсетуі мүмкін.